
Page 54 - 南京医科大学学报自然科学版
P. 54
第42卷第6期
·806 · 南 京 医 科 大 学 学 报 2022年6月
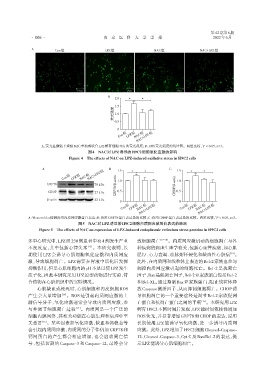
A Con组 LPZ组 NAC组 NAC+LPZ组
B
2.0 * *
1.5
荧光强度 1.0
0.5
0
NAC+LPZ组
Con组 LPZ组 NAC组
A:荧光显微镜下观察NAC单独或联合LPZ孵育细胞48 h的荧光强度;B:ROS荧光强度的统计图。两组比较,P < 0.05,n=3。
*
图4 NAC对LPZ诱导的H9C2细胞氧化应激的影响
Figure 4 The effects of NAC on LPZ⁃induced oxidative stress in H9C2 cells
A B 1.5 * * C 1.5 * *
Con组 LPZ组 NAC组 NAC+LPZ组
GRP78 78 kDa GRP78/β⁃actin 1.0 CHOP/β⁃actin 1.0
CHOP 27 kDa 0.5 0.5
β⁃actin 42 kDa
0 0
NAC+LPZ组
Con组 LPZ组 NAC组 Con组 LPZ组 NAC组
NAC+LPZ组
A:Western blot检测各组内质网应激蛋白表达;B:各组GRP78蛋白表达量的比较;C:各组CHOP蛋白表达量的比较。两组比较,P < 0.05,n=3。
*
图5 NAC对LPZ诱导的H9C2细胞内质网应激蛋白表达的影响
Figure 5 The effects of NAC on expression of LPZ⁃induced endoplasmic reticulum stress proteins in H9C2 cells
多中心研究中,LPZ组230例患者中有4例发生严重 致细胞凋亡 [17-18] 。内质网应激启动的细胞凋亡与各
不良反应,其中包括心律失常 [13] 。本研究表明,长 种疾病的病理生理学有关,包括心血管疾病,如心肌
期使用 LPZ 会诱导心肌细胞氧化应激和内质网应 肥厚、心力衰竭、动脉粥样硬化和缺血性心脏病 。
[19]
激,导致细胞凋亡。LPZ 需要在胃液中活化后发挥 此外,在内质网和线粒体上表达的Bcl⁃2家族也参与
抑酸作用,但是心肌细胞内的pH不足以使LPZ发生 调控内质网应激引起的细胞死亡。Bcl⁃2 是抗凋亡
质子化,因此本研究采用LPZ原型药物进行实验,符 因子,Bax是促凋亡因子,Bcl⁃2亚家族蛋白包括Bcl⁃2
合药物在心脏组织中的实际情况。 和Bcl⁃XL,通过抑制Bax亚家族蛋白,阻止线粒体释
心肌缺血或梗死时,心肌细胞和内皮细胞 ROS 放Caspase激活因子,从而抑制细胞凋亡。CHOP诱
产生会大量增加 [14] 。ROS 是引起内质网应激的上 导细胞凋亡的一个重要途径是调节 Bcl⁃2 家族促凋
游信号分子,氧化应激通常会导致内质网应激,参 亡蛋白和抗凋亡蛋白之间的平衡 [19] 。本研究用LPZ
与并调节细胞凋亡过程 [15] 。内质网是一个广泛的 孵育H9C2不同时间后发现,LPZ能时间依赖性增加
细胞内膜网络,其相关功能在心脏生理和病理中至 ROS荧光,并显著增加GRP78和CHOP的表达,说明
关重要 [16] 。某些因素如氧化应激、缺血和钙稳态等 长期使用 LPZ 能诱导氧化应激,进一步诱导内质网
会引起内质网应激,内质网的分子伴侣如GRP78和 应激。此外,LPZ增加了H9C2细胞Cleaved⁃Caspase⁃
钙网蛋白的产生都会相应增加,也会启动凋亡信 12、Cleaved⁃Caspase⁃3、Cyt C 及Bax/Bcl⁃2的表达,提
号,包括切割的 Caspase⁃3 和 Caspase⁃12,最终会导 示LPZ能诱导心肌细胞凋亡。