
Page 30 - 南京医科大学学报自然科学版
P. 30
第42卷第4期
·480 · 南 京 医 科 大 学 学 报 2022年4月
A B
Control组 PBS+P组 AAV+P组 A20+P组 0.6
( 3D ) ( mm ) *** *** *** ***
腭侧 0.4
( 2D ) 腭侧CEJ⁃ABC的距离 0.2
腭侧 0
Control组 PBS+P组 AAV+P组 A20+P组
C
Control组 PBS+P组 AAV+P组 A20+P组
50
×
100
×
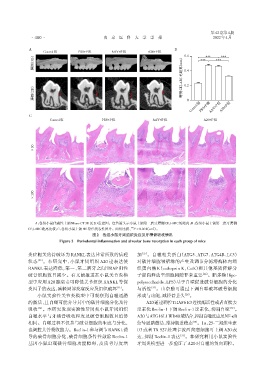
A:各组小鼠代表性上颌Micro⁃CT 3D及2D重建图。红色箭头示小鼠上颌第二磨牙腭侧CEJ⁃ABC的距离;B:各组小鼠上颌第二磨牙腭侧
CEJ⁃ABC距离比较;C:各组小鼠上颌HE染色代表性图片。两组比较, P < 0.001(n=5)。
***
图2 各组小鼠牙周组织炎症及牙槽骨吸收情况
Figure 2 Periodontal inflammation and alveolar bone resorption in each group of mice
炎症相关的骨破坏为 RANKL 表达异常所致的病理 加 [25] 。自噬相关蛋白(ATG5、ATG7、ATG4B、LC3)
状态 [22] 。本研究中,小鼠牙周组织 A20 过表达使 对破骨细胞皱褶缘的产生及调节分泌溶酶体内组
RANKL 表达降低,第一、第二磨牙之间 TRAP 阳性 织蛋白酶 K(cathepsin K,CatK)和其他基质降解分
破骨细胞数目减少。有文献报道在小鼠关节炎模 子定向释放至细胞间隙非常重要 [26] 。脂多糖(lipo⁃
型中应用 A20 腺病毒可降低关节组织 RANKL 等促 polysaccharide,LPS)基于自噬促进破骨细胞的分化
炎因子的表达,减轻局部炎症反应及组织破坏 。 与活性 [27] 。山奈酚可通过下调自噬破坏破骨细胞
[23]
[28]
小鼠实验性关节炎模型中可观察到自噬通路 形成与功能,减轻骨丢失 。
的激活,且自噬可能介导其中的破骨细胞分化及骨 A20通过调控TRAF6 E3连接酶活性或者直接去
吸收 [24] 。本研究发现实验性牙周炎小鼠牙周组织 泛素化Beclin⁃1下调Beclin⁃1泛素化,抑制自噬 [29] 。
自噬水平与牙槽骨吸收程度及破骨细胞数目趋势 A20与ATG16L1 WD40域结合,控制自噬反应及NF⁃κB
相同。自噬过程不仅参与破骨细胞的生成与分化, 信号通路激活,维持肠道稳态 。1α,25⁃二羟维生素
[30]
也调控其骨吸收能力。Beclin⁃1参与调节RANKL诱 D3 或者 TX 527 处理卡波西肉瘤细胞可上调 A20 表
导的破骨细胞分化,破骨细胞条件性敲除 Beclin⁃1 达,抑制 Beclin⁃1 表达 [31] 。本研究利用小鼠实验性
基因小鼠出现破骨细胞功能障碍,皮质骨厚度增 牙周炎模型进一步验证了A20对自噬的负向调控。