
Page 9 - 南京医科大学自然版
P. 9
第45卷第6期 朱淋淋,张 晶,周其冈. 成体海马神经发生与抑郁症:假说与证据[J].
2025年6月 南京医科大学学报(自然科学版),2025,45(6):745-755 ·747 ·
[30-31]
Nestin和GFAP 。 1.3 调控AHN的相关因素
2b 型 NSC 主要通过对称分裂产生 3 型 NSC,也 AHN 受多种因素调控,这些因素共同作用于
可能通过不对称分裂生成。3型NSC表现出神经元 NSC的增殖、分化和成熟过程,进而影响海马的结构
谱系特征,增殖活性较低,具有切向或放射状的突 和功能可塑性。AHN的调控机制复杂,涉及分子信
起 [32] 。这些细胞从SGZ 迁移到GCL,这一迁移过程 号通路、代谢调控、炎症反应、表观遗传修饰以及
通常需要几周甚至数月。在此过程中,3型NSC逐渐 肠⁃脑轴的交互作用等多个层面,这些机制在神经精
分化为海马DG颗粒细胞 。在早期分化阶段,3型 神疾病(如抑郁症)中扮演重要角色。
[33]
NSC 开始表达 DCX,表现出典型的迁移特征。随着 在分子信号通路的调控方面,神经营养因子如
DCX的持续表达,细胞在GCL中进一步成熟,形成树 脑源性神经营养因子(brain⁃derived neurotrophic fac⁃
突和轴突。当细胞开始表达神经元核蛋白(neuronal tor,BDNF)、神 经 生 长 因 子(nerve growth factor,
nuclear protein,NeuN)时,标志着其进入晚期分化阶 NGF)、胶 质 细 胞 源 性 神 经 营 养 因 子(glial cell⁃
段,成为成熟的神经元。此时,细胞展现出丰富的树 derived neurotrophic factor,GDNF)等通过激活细胞
突和轴突结构,并完全整合到海马神经环路中。 内信号通路(如Ras/MAPK、PI3K/Akt、mTOR、Wnt等)
[34]
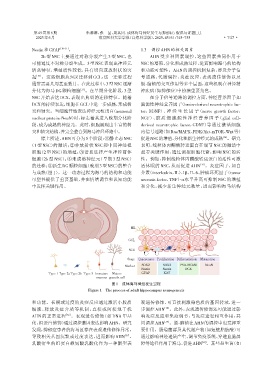
综上所述,AHN可分为5个阶段:①静止态NSC 促进NSC的增殖、分化和新生神经元的成熟 。研究
(1 型 NSC)的激活;②非放射状 NSC 和中间神经祖 表明,线粒体丙酮酸转运蛋白在调节NSC的激活中
细胞(2 型 NSC)的增殖;③谱系选择产生神经前体 起着关键作用,通过调控细胞代谢,影响 NSC 的活
细胞(2b 型 NSC);④未成熟神经元(早期 3 型 NSC) 性。例如,抑制线粒体丙酮酸转运蛋白的活性可激
的迁移;⑤新生DG颗粒细胞(晚期3型NSC)的整合 活休眠的 NSC,从而促进 AHN [35] 。炎症因子,如白
与成熟(图1)。这一动态过程为海马的结构和功能 介素(interleukin,IL)⁃1β、IL⁃6、肿瘤坏死因子(tumor
可塑性提供了重要基础,并在情绪调节和认知功能 necrosis factor,TNF)⁃α水平升高可抑制 NSC 的增殖
中发挥关键作用。 和分化,减少新生神经元数量,进而影响海马结构
ML
GCL
SGZ
Stage Quiescence Proliferation Differentiation Maturation
SOX2 SOX2 PSA⁃NCAM NeuN
Marker
Nestin Nestin DCX
Type 1 Type 2a Type 2b Type 3 Immature Mature GFAP Ki67
neurons granule cell
图1 成体海马神经发生过程
Figure 1 The process of adult hippocampus neurogenesis
和功能。长期或过度的炎症反应通过激活小胶质 观遗传修饰,可直接刺激染色质的基因转录,进一
细胞、释放炎症介质等机制,直接或间接地干扰 步调控AHN [38] 。此外,表观遗传修饰还可能通过影
AHN 的正常进程 [36] 。表观遗传修饰(如 DNA 甲基 响炎症反应和免疫调节,与炎症途径相互作用,共
化、组蛋白修饰)通过调控基因表达影响AHN。研究 同调控AHN [39] 。肠⁃脑轴在AHN的调控中也发挥重
发现,抑郁症患者的海马区存在表观遗传修饰异常, 要作用。肠道菌群及其代谢产物(如短链脂肪酸)可
[37]
导致相关基因沉默或过度表达,进而影响 AHN 。 通过影响神经递质产生、调节免疫系统、穿透血脑屏
乳酸衍生的组蛋白赖氨酸乳酸化作为一种新型表 障等途径作用于海马,促进AHN 。某些益生菌(如
[40]