
Page 136 - 南京医科大学学报自然科学版
P. 136
第41卷第11期
·1698 · 南 京 医 科 大 学 学 报 2021年11月
Micro⁃organisms
β⁃glucan,BCG
Endogenous molecules
oxLDL
Lp(a)
adrenaline
noradrenaline Akt
aldosterone
mTOR
Epigenetic
glucose glucose programs cholesterol
HIF1α
SET7
glycolysis
KDM5 HAT mevalonate
cholesterol
biosynthesis
acetyl⁃CoA
pyruvate
fatty acid
acetyl⁃CoA citrate synthesis
lactate lactate oxaloacetate cis⁃aconitate fatty acids
itaconate
OXPHOS malate TCA cycle
glutamate
α⁃ketoglutarate
NADH glutaminolysis
fumarate
FADH2
succinate glutamine
glutamine glutamine
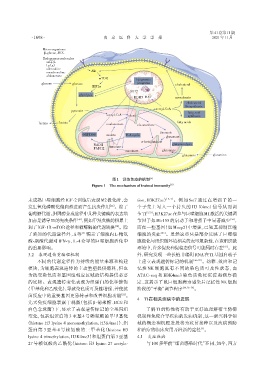
图1 训练免疫的机制 [6]
Figure 1 The mechanism of trained immunity [6]
未成熟巨噬细胞经IGF⁃2训练后表现M2极化时,会 tion,H3K27ac) [3,5] 。例如 Set7 通过在增强子的一
发生氧化磷酸化偏向性进而产生抗炎作用 。除了 个子集上写入一个持久的 H3 K4me1 信号从而调
[23]
葡萄糖代谢,胆固醇合成途径中几种关键酶的表达增 节 TI [5,53] ;H3K27ac在参与巨噬细胞M1激活的关键调
加亦是诱导TI的先决条件 ,例如甲羟戊酸的积累上 节因子如Mir155的启动子和增强子中显著减少 ,
[49]
[54]
[50]
调了IGF⁃1R⁃mTOR途径和糖酵解的代谢转换 。除 而在一些基因(如 Mecp2)中增强,已知其抑制巨噬
了熟知的代谢途径外,Ji 等 [51] 揭示了细胞内L⁃精氨 细胞的炎症 [23] 。虽然这些只是部分反映了巨噬细
酸⁃肌酸代谢对 IFN⁃γ、IL⁃4 介导的巨噬细胞活化中 胞极化与组织微环境相关的表型复杂性,在该组织微
[55]
的重要影响。 环境中,许多促炎和促稳态信号可能同时存在 。此
3.2 表观遗传重编程机制 外,研究发现一些长链非编码RNA在TI基因启动子
不同的代谢途径作为持续的能量来源和构建 上建立表观遗传标记的机制 [56-57] 。幼稚、效应和记
模块,为细胞表观遗传的主动重塑提供燃料,但也 忆性 NK 细胞具有不同的染色质可及性状态,由
为改变染色质和基因组相应区域的结构提供必要 ATAC⁃seq 和 H3K4me3 染色质免疫沉淀和测序确
的底物。表观遗传变化表现为组蛋白的化学修饰 定,这揭示了鼠巨细胞病毒感染后记忆性 NK 细胞
(甲基化和乙酰化),导致染色质可及性增强,并使抗 阶段的“平衡”调节程序 [39,58-59] 。
[52]
菌反应中的重要基因更易转录和改善细胞功能 。
4 TI在相关疾病中的进展
先天免疫细胞暴露于刺激(包括β⁃葡聚糖、BCG 和
白色念珠菌)下,显示了表观遗传标记的全基因组 了解 TI 的特性将有助于更好地理解宿主防御
变化,包括组蛋白第 3 亚基 4 号赖氨酸的单甲基化 机制和免疫介导疾病的发病机制,这一新兴科学领
(histone H3 lysine 4 monomethylation,H3K4me1)、组 域的概念和机理进展将为疫苗接种以及疾病预防
[3]
蛋白第 3 亚基 4 号赖氨酸的三甲基化(histone H3 和治疗的临床应用开辟新的途径 。
lysine 4 trimethylation,H3K4me3)和组蛋白第 3 亚基 4.1 炎症疾病
27 号赖氨酸的乙酰化(histone H3 lysine 27 acetyla⁃ 与100多年前“细菌感染时代”不同,如今,西方