
Page 34 - 南京医科大学学报自然科学版
P. 34
第42卷第5期
·630 · 南 京 医 科 大 学 学 报 2022年5月
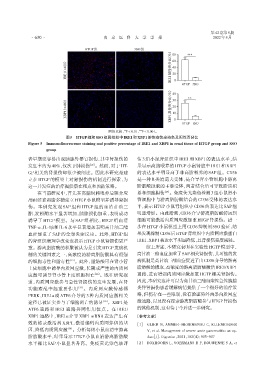
HTGP组 SSO组
( % ) 80 ***
( ×400 ) IRE1阳性百分比 60
40
IRE1 20
0
HTGP SSO
( % ) 40 **
( ×400 ) XBP1阳性百分比 30
20
XBP1 10
0
HTGP SSO
两组比较,Р < 0.01, Р < 0.001。
***
**
图5 HTGP组和SSO组肾组织中IRE1和XBP1的免疫荧光染色及阳性百分比
Figure 5 Immunofluorescence staining and positive percentage of IRE1 and XBP1 in renal tissue of HTGP group and SSO
group
者早期更容易出现胰腺外器官损伤,其中肾损伤的 估 3 组小鼠肾组织中 IRE1 和 XBP1 的表达水平,结
发生率约为40%,仅次于肺损伤 [25] 。然而,对于HT⁃ 果显示高脂喂养的HTGP 小鼠肾脏中IRE1和XBP1
GP 相关的肾损伤却很少被阐述。因此本研究是建 的表达水平明显高于非高脂喂养的 SAP 组。CD36
立在 HTGP 的模型上对肾损伤的机制进行探索,为 是一种 B 类清道夫受体,是介导肾小管细胞中游离
这一并发症的治疗提供潜在靶点和预防策略。 脂肪酸摄取的主要受体,两者结合后可导致脂质积
在当前研究中,首先在高脂饲料喂养后联合应 累和细胞损伤 [29] 。免疫荧光染色检测3组小鼠肾小
用雨蛙素和脂多糖建立HTGP小鼠模型并诱导肾损 管细胞中与游离脂肪酸结合的CD36受体的表达水
伤。本研究发现 SAP 组和 HTGP 组的血清甘油三 平,提示HTGP小鼠肾组织中CD36的表达比SAP组
酯、淀粉酶水平显著增加,胰腺损伤加重,表明成功 明显增加。由此猜测,CD36介导游离脂肪酸转运到
诱导了 HTGP 模型。与 SAP 组相比,HTGP 组血清 细胞可能激活内质网应激加重HTGP肾损伤。进一
TNF⁃α、IL⁃1β和IL⁃6水平显著增加表明高甘油三酯 步在HTGP小鼠模型上用CD36抑制剂SSO验证,结
血症加重了 SAP 的全身炎症反应。此外,HTGP 组 果发现抑制CD36后HTGP肾组织中内质网应激蛋白
的肾组织病理学改变也提示HTGP小鼠肾损伤更严 IRE1、XBP1表达水平明显降低,且肾损伤程度减轻。
重。游离脂肪酸的积聚被认为是引发HTGP发病机 综上所述,本研究表明在实验性HGTP模型中,
制的关键因素之一,高浓度的游离脂肪酸具有很强 高甘油三酯血症加重了SAP相关肾损伤,其可能的发
的细胞毒性和脂毒性 [13] 。此外,脂肪酸可在肾小管 病机制是高甘油三酯血症促进了由CD36介导的游离
上皮细胞中诱导内质网应激,长期或严重的内质网 脂肪酸的摄取,高浓度的游离脂肪酸激活IRE1/XBP1
应激可诱导肾小管上皮细胞凋亡 [26] 。既往研究报 通路,进而增强内质网应激加重HGTP相关肾损伤。
道,内质网应激参与急性肾损伤的发生发展,在肾 因此,本研究也许可以为高甘油三酯血症性急性胰腺
功能紊乱中起重要作用 [27] 。内质网应激传感器 炎伴肾损伤患者缓解病情提供了一个很好的治疗策
PERK、IRE1α或 ATF6 介导的 3 种内质网应激相关 略,但仍存在一些限制,没有验证另外两条内质网应
途径已被证实参与了细胞凋亡的诱导 [10] 。XBP1 是 激通路,以及没有探索游离脂肪酸参与HTGP肾损伤
的其他机制,这有待于今后进一步研究。
ATF6 通路和 IRE1 通路共同作用位点。在 IRE1/
XBP1 通路中,IRE1α介导 XBP1 mRNA 表达产生有 [参考文献]
效的转录激活剂 XBP1,激活编码内质网伴侣的基 [1] GLIEM N,AMMER⁃HERRMENAU C,ELLENRIEDER
因,降低内质网应激 [28] 。分析每组小鼠血清中游离 V,et al. Management of severe acute pancreatitis:an up⁃
脂肪酸水平,结果显示 HTGP 小鼠血清游离脂肪酸 date[J]. Digestion,2021,102(4):503-507
水平相比 SAP 小鼠显著升高。免疫荧光染色法评 [2] BOXHOORN L,VOERMANS R P,BOUWENSE S A,et