
Page 65 - 南京医科大学学报自然科学版
P. 65
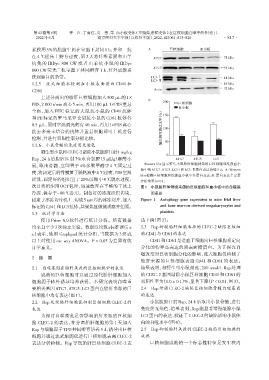
第42卷第6期 姜 江,丁育红,贾 慧,等. 血小板受体C型凝集素样受体2在巨核细胞自噬中的作用[J].
2022年6月 南京医科大学学报(自然科学版),2022,42(06):815-820 ·817 ·
素膜用 5%的脱脂牛奶在室温下封闭 1 h,并和一抗 A 巨核细胞 血小板
在 4 ℃摇床上孵育过夜,第 2 天将纤维素膜和山羊
ATG7 ←78 kDa
抗兔的 IRDye 800 CW 或者山羊抗小鼠的 IRDye
800 CW荧光二抗室温下共同孵育 1 h,红外成像系 ATG5 ←55 kDa
统观察目的条带。 LC3⁃Ⅰ ←14 kDa
LC3⁃Ⅱ
1.2.5 流式细胞术检测血小板表面蛋白 CD41 和 ←16 kDa
←42 kDa
CD61 Actin
上述分离出的胎肝巨核细胞加入 500 μL 的 1×
B 200
PBS,2 000 r/min离心5 min,再用100 μL 1×PBS 重悬 巨核细胞
血小板
全血,加入 FITC 标记的大鼠抗小鼠的 CD41 抗体
和 PE 标记的罗马尼亚仓鼠抗小鼠的 CD61 抗体各 ( % ) 150
0.5 μL,同时室温避光孵育40 min,再用1×PBS离心
洗去多余未结合的抗体并重悬细胞即可上机进行 蛋白相对表达水平 100
检测,并进行双阳性率分析比较。
1.2.6 小鼠骨髓的免疫荧光染色 50
野生型小鼠和CLEC⁃2敲除小鼠腹腔注射1 mg/kg
0
Rap,24 h 后腹腔注射 7%水合氯醛(5 μL/g)麻醉小 ATG7 ATG5 LC3
鼠,取出骨髓,立即置于 4%多聚甲醛中 4 ℃固定过 Western blot 显示野生小鼠胎肝细胞诱导的巨核细胞和洗涤血小
板中的 ATG7、ATG5、LC3⁃I 和 LC3⁃Ⅱ蛋白表达持续 5 d。A:Western
夜,将固定后的骨髓置于脱钙液中4 ℃过夜,PBS室温
blot 检测巨核细胞和洗涤血小板中各蛋白表达;B:蛋白表达半定量
摇洗,固定好的组织置于20%蔗糖中4 ℃脱水过夜, 分析结果(n=3)。
次日将组织用OCT包埋,迅速放置在平整的干冰上 图1 小鼠胎肝和骨髓来源的巨核细胞和血小板中的自噬基
冷冻,保存于-80 ℃备用。制备好的冰冻组织切块, 因表达
固定于冰冻切片机上,切成5 μm厚的冰冻切片,加入 Figure 1 Autophagy gene expression in mice fetal liver
标记的CD41和LC3抗体,共聚焦显微镜观察并拍照。 and bone marrow⁃derived megakaryocytes and
platelets
1.3 统计学方法
使用 Prism 6.0 软件进行统计分析。所有数据 达下调(图2)。
均来自至少3次独立实验。数据以均数±标准误(x ± 2.3 Rap 抑制胎肝细胞来源的 CLEC⁃2 缺陷巨核细
sx )表示,使用 Graphpad 统计分析,当数据为 3 组或 胞CD41与CD61的表达
以上时使用 one way ANOVA。P < 0.05 为差异有统 CD41和CD61是造血干细胞向巨核细胞系定向
计学意义。 分化的特异高表达的膜表面糖蛋白。为了探究自
噬改变对巨核细胞分化的影响,流式细胞仪检测了
2 结 果
胎肝来源的巨核细胞表面 CD41 和 CD61 的表达。
2.1 自噬基因在胎肝来源的巨核细胞中的表达 结果表明,和野生型小鼠相比,200 nmol/L Rap 处理
成熟的巨核细胞可以通过原代胎肝细胞加入 的CLEC⁃2基因敲除小鼠巨核细胞CD41和CD61的
细胞因子体外诱导培养获得。本研究表明自噬重 双阳性率为(1.0 ± 0.1)%,显著下降(P < 0.01,图3)。
要相关基因ATG7、ATG5、LC3蛋白在胎肝来源的巨 2.4 Rap 增强 CLEC⁃2 缺陷巨核细胞骨髓自噬基因
核细胞中均有表达(图1)。 的表达
2.2 Rap 处理胎肝细胞能抑制巨核细胞 CLEC⁃2 的 小鼠腹腔注射Rap,24 h后取出小鼠骨髓,进行
表达 免疫荧光染色,结果表明,Rap能显著增强敲除小鼠
为探讨自噬紊乱是否影响胎肝来源的巨核细 LC3蛋白的表达,验证了CLEC⁃2的缺陷影响小鼠体
胞 CLEC⁃2 的表达,在分离胎肝细胞的第 1 天加入 内的自噬水平(图4)。
Rap与细胞因子TPO共同孵育培养5 d,诱导出巨核 2.5 Rap 抑制胎肝来源的 CLEC⁃2 缺陷巨核细胞的
细胞并通过流式细胞仪进行巨核细胞表面 CLEC⁃2 成熟
表达分析检测。Rap 导致胎肝巨核细胞 CLEC⁃2 表 巨核细胞成熟的一个标志性特征是发生核内