
Page 42 - 南京医科大学学报自然科学版
P. 42
第42卷第7期
·938 · 南 京 医 科 大 学 学 报 2022年7月
A 多西环素饮食 B C
CK CK/ALCAT1 KO □ CK □ CK
1.5 ■ CK/ALCAT1 KO 1.5 ■ CK/ALCAT1 KO
p⁃Akt(T308) 60 kDa
**
/Akt 1.0 1.0
Akt 60 kDa
( T308 )
p⁃Erk1/2 44/42 kDa p⁃Erk1/2/Erk1/2
p⁃Akt 0.5 0.5
44/42 kDa
Erk1/2
37 kDa 0 0
GAPDH
多西环素饮食 多西环素饮食
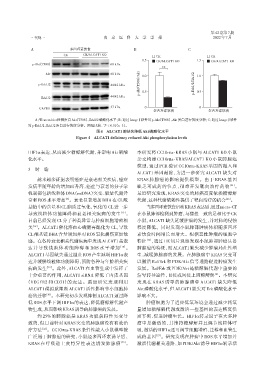
A:Western blot检测蛋白Akt(T308)、Erk1/2磷酸化水平;B:通过Image J软件对p⁃Akt(T308)、Akt蛋白进行灰度分析;C:通过Image J软件
对p⁃Erk1/2、Erk1/2蛋白进行灰度分析。两组比较,P < 0.01(n=3)。
**
图4 ALCAT1的缺失降低Akt磷酸化水平
Figure 4 ALCAT1 deficiency reduced Akt phosphorylation levels
HIF1α表达,从而减少糖酵解代谢,并影响 Akt 磷酸 本研究将 CC10rtta⁃KRAS 小鼠与 ALCAT1 KO 小鼠
化水平。 杂交构建 CC10rtta⁃KRAS/ALCAT1 KO 小鼠肺腺癌
模型,通过 PCR 验证 CC10rtta⁃KRAS 基因的敲入和
3 讨 论
ALCAT1 基因敲除,为进一步研究 ALCAT1 缺失对
越来越多证据表明癌症是衰老相关疾病,癌症 KRAS 肺腺癌的影响提供模型。由于 KRAS 基因
发病率随年龄的增加而升高,癌症与衰老的分子学 缺乏可成药的位点,很难开发靶向治疗药物 [24] 。
机制都包括线粒体DNA(mtDNA)突变、能量代谢异 最近研究发现,KRAS 突变的肺癌高度依赖糖酵解
[9]
[25]
常和ROS水平增加 。衰老显著增加DHA在CL酰 代谢,这种代谢依赖性提供了靶向治疗的机会 。
基链中的含量和CL 脂质过氧化,氧化的 CL 进一步 当多西环素饮食启动KRAS表达后,通过micro⁃CT
导致线粒体功能障碍和衰老相关疾病的发生 [13] 。 在小鼠肺部检测到肿瘤,与猜想一致的是相比于CK
目前已经发现 CL 分子种类异常与肿瘤细胞增殖相 小鼠,ALCAT1缺失延缓肿瘤的发生,并对肺的侵蚀
关 [16] 。ALCAT1催化溶血心磷脂再酰化为 CL,导致 程度降低。同时发现小鼠肺部肿瘤体积随多西环
CL酰基链DHA含量增加并对ROS氧化损伤更加敏 素饮食时间增长而增大。根据恶性肿瘤的细胞学
感。在各种衰老相关代谢疾病中发现ALCAT1高表 特征 [26] ,通过 HE 切片观察发现小鼠肺部肿瘤具备
达并导致线粒体功能障碍和 ROS 水平增加 [18] 。 肺腺癌的特征,而 ALCAT1 缺失减少肿瘤局灶性增
ALCAT1基因缺失通过阻止ROS产生减弱HIF1α表 生,减缓肺腺癌的发展。在肺腺癌中 KRAS 突变可
达并缓解线粒体功能障碍,预防各种与年龄相关疾 以激活 Raf/Erk 和 PI3K/Akt 信号通路促进肿瘤发生
病的发生 [19] 。此外,ALCAT1 在血管生成中发挥了 发展。Raf/Erk 或 PI3K/Akt 是糖酵解代谢中重要的
十分重要的作用,ALCAT1 siRNA 降低了内皮基因 信号转导途径,直接或间接上调糖酵解 。本研究
[4]
(VEGFR2 和 CD31)的表达。最近研究发现利用 发现在 KRAS 诱导的肺腺癌中 ALCAT1 缺失降低
ALCAT1 模拟肽阻断 ALCAT1 活性影响非小细胞肺 Akt磷酸化水平,但ALCAT1缺失对Erk磷酸化水平
癌的迁移 [20] 。本研究初步发现抑制ALCAT1通过降 影响不大。
低 ROS 水平下调 HIF1α的表达,降低糖酵解代谢产 肿瘤细胞为了适应低氧环境会通过减少耗氧
物生成,从而影响KRAS诱导的肺腺癌的发展。 量增加糖酵解代谢或激活一些基因的表达恢复供
约 25%的肺腺癌是 KRAS 功能获得性突变导 需平衡、促进肿瘤生长。HIF1α转录因子在大多肿
致的,但目前针对 KRAS 突变的肺腺癌没有有效治 瘤中是激活的,其维持糖酵解并且调节线粒体呼
疗方法 [22] 。CC10rtta⁃KRAS 条件性敲入小鼠模型被 吸,激活的HIF1α还可调节细胞增殖、迁移和血管生
广泛用于肺腺癌的研究,小鼠经多西环素诱导后, 成的基因 [27] 。研究发现在肿瘤中 ROS 水平增加并
KRAS 在呼吸道上皮特异性表达诱发肺腺癌 [23] 。 激活代谢相关通路,如 PI3K/Akt 诱导 HIF1α转录活