
Page 36 - 南京医科大学自然版
P. 36
第45卷第3期
·324 · 南 京 医 科 大 学 学 报 2025年3月
A 0 h 48 h 72 h
B C
6 5 2 2 10 8
Virus titer (lgTCID50/mL) 4 3 2 HA titer 2 2 6 4
0 1 2 2 2 0
0 12 24 36 48 60 72 0 24 48 72 96 120 144
Time post infection(h) Time post infection(h)
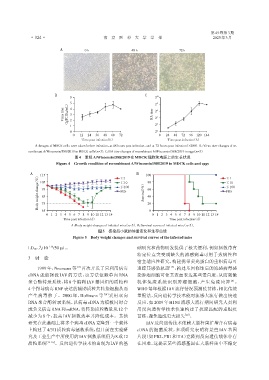
A:Images of MDCK cells were taken before infection,at 48 hours post infection,and at 72 hours post infection(×200). B:Virus titer changes of re⁃
combinant A/Wisconsin/588/2019 in MDCK cells(n=3). C:HA titer changes of recombinant A/Wisconsin/588/2019 in eggs(n=3).
图4 重组A/Wisconsin/588/2019在MDCK细胞和鸡胚上的生长状况
Figure 4 Growth condition of recombinant A/Wisconsin/588/2019 in MDCK cells and eggs
A 115 B 100
(%) 105 1∶1 (%) 80 1∶1
1∶10
1∶10
Body weight change 95 PBS Survival 60 PBS
1∶100
1∶100
85
40
20
75
65 0
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 0 1 2 3 4 5 6 7 8 9 10 11 12 13 14
Time post infection(d) Time post infection(d)
A:Body weight changes of infected mice(n=5). B:Survival curves of infected mice(n=5).
图5 感染后小鼠的体重变化和生存曲线
Figure 5 Body weight changes and survival curves of the infected mice
LD50,为10 -1.38 /50 μL。 础研究和药物研发提供了极大便利,例如拯救带有
特定位点突变或缺失的流感病毒可用于致病性和
3 讨 论
宿主适应性研究,构建携带荧光蛋白的重组病毒可
1999 年,Neumann 等 [13] 首次开发了只利用病毒 追踪其感染轨迹 [17] ,构建基因修饰后的流感病毒感
cDNA 就能拯救 IAV 的方法,该方法依赖单向 RNA 染肿瘤细胞可使其表面表达某些蛋白质,从而刺激
[8]
聚合酶转录系统,将 8 个编码 IAV 基因组的质粒和 机体免疫系统识别肿瘤细胞,产生免疫应答 。
4 个指导病毒RNP表达的辅助质粒共转染细胞从而 WHO每年根据IAV流行情况预测疫苗株,相比传统
产生病毒粒子。2000 年,Hoffmann 等 [14] 采用双向 重配法,反向遗传学技术能对流感大流行做出快速
RNA 聚合酶转录系统,以病毒 cDNA 为模板同时合 反应,如 2009 年 H1N1 流感大流行期间研究人员利
成负义病毒RNA和mRNA,将转染质粒数量从12个 用反向遗传学技术快速构建了抗原匹配的重组疫
[10]
减少为 8 个,提高 IAV 拯救效率并降低成本。其他 苗株,避免造成更大损失 。
研究在此基础上将多个病毒cDNA克隆到一个载体 IAV反向遗传技术依赖大肠杆菌扩增含有病毒
上构建了4/3/1质粒病毒拯救系统,但目前在实验研 cDNA 的细菌质粒,多项研究表明将某些 IAV 基因
究及工业生产中所使用的IAV拯救系统仍为8或12 片段(如PB2、PB1和HA)克隆到反向遗传载体中存
质粒系统 [14-16] 。反向遗传学技术的出现为IAV的基 在困难,这提示某些流感基因在大肠杆菌中不稳定